VON MARTIN A. BILLETER
Impfstoffe zur Verhinderung von Infektionskrankheiten, seien sie durch Kleinlebewesen oder durch Viren hervorgerufen, sind nicht mehr aus dem Repertoire medizinischer Wirkstoffe wegzudenken. Der wohl spektakulärste Erfolg von systematischen Impfkampagnen ist die weltweite Ausrottung der Pocken. Auf der einen Seite wird an der Neuentwicklung von Impfstoffen intensiv geforscht, auf der anderen warnen Impfgegner vor Fehlentwicklungen oder stellen den Nutzen von Impfungen generell in Frage. Hier einige Erläuterungen zur Wirkungsweise und Entwicklung von Impfstoffen.
[ Diagramm ]
Das Prinzip aller Impfungen ist sehr einfach. Dem Individuum, das vorbeugend gegen eine Infektionskrankheit geschützt werden soll, werden abgeschwächte Varianten des Krankheitserregers selbst (Lebendimpfstoffe) oder aber abgetötete Krankheitserreger (Totimpfstoffe) appliziert; anstelle der letzteren gelangen heutzutage häufig Einzelbestandteile davon zum Einsatz (Komponentenimpfstoffe), welche oft gentechnisch hergestellt sind. Impfstoffe regen das Immunsystem zur Bildung von Abwehr-Agentien, das heisst Antikörpern und Immunzellen mitsamt ihren Wirkstoffen, an, so dass im Falle einer Infektion der entsprechende Erreger unverzüglich bekämpft wird. Anders gesagt, das Immunsystem, welches normalerweise in jedem Individuum auf Infektionen und andere Abnormalitäten (zum Beispiel Krebszellen) mit einer gewissen Verzögerung reagiert, soll präventiv zur Bildung eines erregerspezifischen Immungedächtnisses stimuliert werden. Dieäusserst komplexe Natur der Immunantwort braucht uns hier nur insofern zu interessieren, als es ausschlaggebend ist, in welcher Form und an welchen Stellen im Körper Impfstoffe appliziert werden. Generell gilt, dass sich wirkungsvolle und lange (manchmal lebenslänglich) andauernde spezifische Immunantworten dann bilden, wenn die Immunstimulation so natürlich wie möglich geschieht. Daher haben sich abgeschwächte (attenuierte) Krankheitserreger, die ihre Virulenz weitgehend verloren haben, beispielsweise weil sie sich nur langsam vermehren oder keine Toxine mehr produzieren, meist alsäusserst wirksam erwiesen. Ideal ist deren Applikationüber den normalen Infektionsweg, zum Beispiel die Atmungswege oder den Magen-Darm-Trakt. Dadurch, dass ein solcher Lebendimpfstoff sich vermehrt, werden seine Bestandteile dem Immunsystem auch an der Oberfläche von infizierten Zellen präsentiert; gewisse Zweige der Immunantwort sindüberhaupt nur auf diese Weise stimulierbar. Ein anderer Vorteil der Lebendimpfstoffe besteht darin, dass dank der Vermehrungsfähigkeit nur sehr geringe Mengen benötigt werden, so dass die Herstellungskosten typischerweise um Zehnerpotenzen niedriger liegen als für Tot- oder Komponentenimpfstoffe. Vor allem für den Grosseinsatz in Entwicklungsländern ist dies von entscheidender Bedeutung. Allerdings sind Lebendimpfstoffe meist hitzelabil, so dass sie in Kühlschränken aufbewahrt werden müssen.
Den grossen Vorteilen steht die zumindest potentielle Gefährlichkeit von Lebendimpfstoffen gegenüber. Der zur Immunisierung gegen Pocken verwendete Vaccinia-Impfstamm löste regelmässig Krankheitserscheinungen aus; obwohl diese meistens eher geringfügige Natur waren, blieben auch schwere Komplikationen nicht aus, die zum Teil tödlich verliefen. Wie ist heute die Gefährlichkeit gebräuchlicher Lebendimpfstoffe einzustufen?
Die Vorbedingung einer niedrigen Virulenz beruht oft darauf, dass der Krankheitserreger an einen anderen Wirt adaptiert ist. Bei der ersten systematischen Impfung, vor rund 200 Jahren, verwendete Jenner Kuhpockenvirus, welches sich im Menschen langsamer vermehrt; da anderseits Kuhpockenvirus und die heute gängigen Laborstämme des verwandten Vacciniavirus dem menschlichen Pockenvirus (Variola) sehrähneln, wirkt das induzierte Immungedächtnis auch sehr gut gegen Variola. Bereits in der ersten Hälfte unseres Jahrhunderts wurde damit begonnen, attenuierte Krankheitserreger durch Mutagenese, das heisst durch ungezielte Veränderung ihrer Erbsubstanz, zu gewinnen mittels Einwirkung von Chemikalien oder verschiedenartiger Strahlung. Dieüberlebenden Erreger, welche zufälligerweise ihre Virulenz verloren hatten, kamen als Lebendimpfstoffe in Frage. Bei der Auswahl galt es zu beachten, dass genügend, aber nicht zuviel der Virulenz verlorengegangen war, denn allzusehr abgeschwächte Erreger induzieren meist auch eine ungenügende Immunantwort. Als Beispiel sei der in der Praxis sehr bewährte Impfstoff gegen Tuberkulose (Bacillus Calmette GuŽrin, BCG) genannt.
Eine interessante Variante dieses Vorgehens besteht darin, spontan entstehende Mutanten zu selektionieren. Wenn Viren während vieler Generationen in einem anderen Wirt vermehrt werden, beispielsweise Viren des Menschen auf Hühner-Embryonen, werden automatisch Virusvarianten selektioniert, welche in ihrer Erbsubstanz Veränderungen (Mutationen) aufweisen, die sie befähigen, sich im neuen Wirt besonders effizient zu vermehren. Solche an den neuen Wirt adaptierte Viren sind dann oft für den alten Wirt viel weniger virulent. Diese Methode ist hauptsächlich bei RNA-Viren anwendbar, die als Erbsubstanz nicht DNA, die Erbsubstanz aller Lebewesen, besitzen, sondern RNA. (RNA-Moleküle stellen normalerweise sozusagen Gebrauchskopien einzelner Regionen der Erbsubstanz DNA dar. In Lebewesen ist RNA ein kurzlebiger Informationsträger, welcher Sequenzinformation, die in der DNA enthalten ist, chemisch verfügbar macht zur Synthese von Proteinen). Bei der Vermehrung dieser Viren wird RNA von RNA abkopiert, ein etwas ungenauer Vorgang, da dabei im Gegensatz zur DNA-Vermehrung keine Korrektur falsch eingebauter Basen stattfindet. Aus diesem Grund erfolgt die spontane Adaptierung an einen neuen Wirt und mithin oft auch die Attenuation gegenüber dem alten besonders rasch. Bereits vor rund 60 Jahren wurde auf diese Weise ein Impfstoff gegen das gefürchtete Gelbfieber gewonnen. Bekannte weitere Beispiele sind die vor beinahe 40 Jahren entwickelten Lebendimpfstoffe gegen Poliomyelitis (Kinderlähmung) und gegen Masern.
Vor rund zwanzig Jahren ist schliesslich die DNA-Klontechnik entwickelt worden, welche es erlaubt, relativ kleine, aus der Natur entnommene oder auch rein chemisch synthetisierte DNA-Segmente miteinander zu kombinieren und in Bakterien zu vermehren. Seither sind viele verschiedenartige Strategien ausgearbeitet worden, um die Erbsubstanz von Bakterien oder Viren, die zum Teil bereits in attenuierter Form als Lebendimpfstoffe erfolgreich in der Praxis eingesetzt waren, gezielt und planmässig zu verändern. So ist es einerseits möglich, Gene, welche für Virulenzfaktoren kodieren, ganz zu eliminieren oder in geeigneter Weise zu verändern, um die Impfstoffe sicherer zu machen. Anderseits können zusätzliche, anderen Krankheitserregern entnommene Gene eingeführt werden; solche rekombinante multivalente Impfstoffe induzieren im Geimpften zusätzliche Immunantworten auch gegenüber den entsprechenden anderen Krankheitserregern. Solche rekombinanten Derivate sind beispielweise von Cholera- und Salmonellenbakterien, Vacciniaviren sowie Adenoviren gewonnen worden; letztere werden auch für somatische Gentherapie eingesetzt. Um solche Manipulationen an den viel kleineren RNA-Viren durchführen zu können, muss deren Erbsubstanz RNA zunächst in DNA umkopiert werden. Nach dem Einbringen der beabsichtigten Veränderungen in die DNA wird daraus wieder RNA abkopiert. Im Falle von vielen einfachen RNA-Viren ist solche abkopierte RNA, wenn sie künstlich in gezüchtete Zellen eingeschleust wird, infektiös, das heisst, es bilden sich Nachkommenviren. Seit rund einem Dutzend Jahren können beispielsweise die oben erwähnten Polioviren bzw. deren attenuierten Abkömmlinge so manipuliert werden. Dies war bislang nicht der Fall für eine grosse Gruppe von Negativ-Strang-Viren, wie beispielsweise Masern- oder Tollwut-Virus, weil deren Erbsubstanz nur aktiv wird, wenn sie in viralen Proteinen verpackt als Ribonukleoprotein (RNP) vorliegt (siehe Abbildung und Erläuterungen unten). Nachdem dieses technische Problem jetzt aber gelöst ist, erweist sich die Art der Verpackung und der Reproduktion der Erbsubstanz bei diesen Viren nun als Vorteil: Es hat sich herausgestellt, dass die oben erwähnte allgemeine Fehleranfälligkeit der RNA-Viren bei dieser Gruppe bedeutend geringer ist. Da diese Viren spontan praktisch nicht rekombinieren (auch aus Sicherheitsgründen ein grosser Vorteil), werden eingesetzte Fremdgene in der Nachkommenschaft nicht eliminiert, wie bei Poliovirus, sondern höchstens punktuell langsam verändert.
Generell sind für Lebendimpfstoffe, wie eingangs erwähnt, die Effizienz hoch und die Produktionskosten niedrig; seit gezielte genetische Veränderungen möglich sind, ist auch das Gefahrenpotential ständig am Sinken. Die Sicherheitsanforderungen, die von den nationalen Gesundheitsbehörden heutzutage gestellt werden können, sind mithin auch bedeutend rigoroser als zu Beginn der verschiedenen systematischen Impfkampagnen. Beispielsweise sind heute multivalente rekombinante Vaccinia-Impfviren für den menschlichen Gebrauch generell nicht zugelassen, trotz des Erfolges der viel virulenteren Vaccinia-Stämme bei der Ausrottung der Pockenseuche. Abschliessend noch eine generelle Bemerkung zur Abschätzung von Gefahrenpotentialen. Intuitiv würde man annehmen, dass die teuren und generell weniger effizienten Tot- bzw. Komponentenimpfstoffe sicherer sind als vermehrungsfähige. Bei Masern war jedoch das Gegenteil der Fall: Abgetötete Masernviren haben eine aberrante Immunantwort zur Folge gehabt, welche die Geimpften nicht nur mangelhaft schützte, sondern bei Infektion eine viel gravierendere Krankheit, atypische Masern, bewirkte, welche auch hierzulande zu etlichen Toten führte (weltweit fordert Masern immer noch rund eine Million Todesopfer pro Jahr, hauptsächlich in Entwicklungsländern; vor der Anwendung der attenuierten Impfstoffe waren es rund dreimal so viele). Für die Nutzen/Schaden-Abschätzung von Impfstoffen gibt es somit nur allgemeine Leitlinien, nicht aber feste Regeln; von Fall zu Fall muss unter Berücksichtigung vieler verschiedener Kriterienüber die Verwendung von vermehrungsfähigen oder toten Impfstoffen entschieden werden. Entscheidungen dieser Art sind auch für die jetzt in Erprobung stehenden DNA-Impfstoffe zu treffen, welche darauf beruhen, Erbsubstanz mit Information zur Herstellung einzelner Proteine direkt in Körperzellen von Impflingen einzuschleusen.
Der Infektionszyklus von Masernvirus (MV)
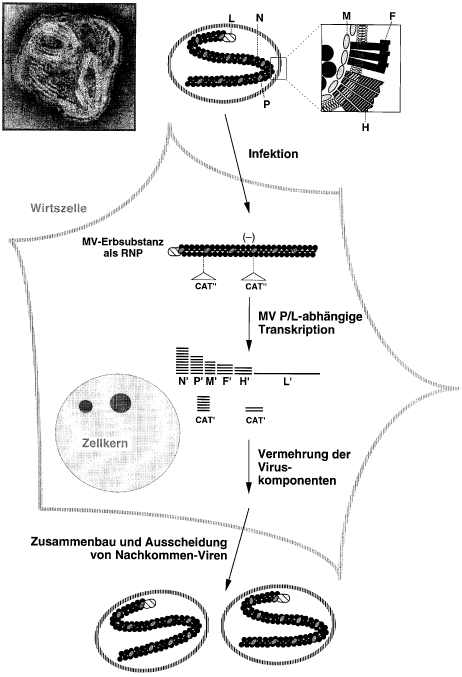 Der obere Teil zeigt eine elektronenmikroskopische Aufnahme sowie ein Schema von MV, welches die Anordnung der sechs verschiedenartigen Virus-Protein-Komponenten andeutet. N, P und L, dicht assoziiert mit der Erbsubstanz RNA (15894 Basen Länge, repräsentiert als dünner Strich), bilden ein Ribonukleoprotein (RNP). M, F und H sind in die umhüllende Membran integriert, die das Virus jeweils von der Membran seiner Wirtszelle usurpiert. Nach Verschmelzen der Virusmembran mit der Zellmembran werden vom RNP zunächst die mit N', P', M', F', H' und L' bezeichneten «RNA-Gebrauchskopien» abgelesen (Transkription), welche zur Herstellung der entsprechenden Proteine benötigt werden. Um MV genetisch verändern zu können, wurde in unserer Arbeitsgruppe einerseits ein in Bakterien vermehrbares etwa 20000 Basen langes DNA-Molekül («MV-DNA») so konstruiert, dass T7 RNA-Polymerase es als Matrize gebrauchen kann, um genaue Kopien der MV-Erbsubstanz künstlich herzustellen. Anderseits wurden in Kulturzellen Gene so integriert, dass die Zellen vier Proteine bilden, und zwar T7 RNA-Polymerase sowie die MV-Proteine N, P und L im richtigen Verhältnis. Wenn in diese Helferzellen «MV-DNA»-Moleküle eingeschleust werden, bildet sich MV-RNA, welche sogleich als RNP verpackt wird; von hier aus setzt sich der Infektionszyklus spontan fort. Fremdgene können nun in die «MV-DNA» eingesetzt werden, wobei die vom Virus vorgegebenen Interpunktionsregeln (Kontrollsignale in Form von Basensequenzen) zu beachten sind. Die daraus abgeleiteten überlangen RNPs führen zu MV-Varianten, welche zusätzliche Proteine, beispielsweise das einfach nachweisbare CAT-Enzym bilden. Die Abbildung zeigt schematisch, an welchen Stellen in den RNPs die bis jetzt eingesetzten Fremdgene lokalisiert sind (Dreiecke, CAT'') und deutet auch an, dass entsprechend ihrer Lokalisation im RNP unterschiedliche Mengen von «RNA-Gebrauchskopien» (CAT') gebildet werden.
Der obere Teil zeigt eine elektronenmikroskopische Aufnahme sowie ein Schema von MV, welches die Anordnung der sechs verschiedenartigen Virus-Protein-Komponenten andeutet. N, P und L, dicht assoziiert mit der Erbsubstanz RNA (15894 Basen Länge, repräsentiert als dünner Strich), bilden ein Ribonukleoprotein (RNP). M, F und H sind in die umhüllende Membran integriert, die das Virus jeweils von der Membran seiner Wirtszelle usurpiert. Nach Verschmelzen der Virusmembran mit der Zellmembran werden vom RNP zunächst die mit N', P', M', F', H' und L' bezeichneten «RNA-Gebrauchskopien» abgelesen (Transkription), welche zur Herstellung der entsprechenden Proteine benötigt werden. Um MV genetisch verändern zu können, wurde in unserer Arbeitsgruppe einerseits ein in Bakterien vermehrbares etwa 20000 Basen langes DNA-Molekül («MV-DNA») so konstruiert, dass T7 RNA-Polymerase es als Matrize gebrauchen kann, um genaue Kopien der MV-Erbsubstanz künstlich herzustellen. Anderseits wurden in Kulturzellen Gene so integriert, dass die Zellen vier Proteine bilden, und zwar T7 RNA-Polymerase sowie die MV-Proteine N, P und L im richtigen Verhältnis. Wenn in diese Helferzellen «MV-DNA»-Moleküle eingeschleust werden, bildet sich MV-RNA, welche sogleich als RNP verpackt wird; von hier aus setzt sich der Infektionszyklus spontan fort. Fremdgene können nun in die «MV-DNA» eingesetzt werden, wobei die vom Virus vorgegebenen Interpunktionsregeln (Kontrollsignale in Form von Basensequenzen) zu beachten sind. Die daraus abgeleiteten überlangen RNPs führen zu MV-Varianten, welche zusätzliche Proteine, beispielsweise das einfach nachweisbare CAT-Enzym bilden. Die Abbildung zeigt schematisch, an welchen Stellen in den RNPs die bis jetzt eingesetzten Fremdgene lokalisiert sind (Dreiecke, CAT'') und deutet auch an, dass entsprechend ihrer Lokalisation im RNP unterschiedliche Mengen von «RNA-Gebrauchskopien» (CAT') gebildet werden.
Dr. Martin A. Billeter (billeter@molbio1.unizh.ch) ist ordentlicher Professor am Institut für Molekularbiologie der Universität Zürich.
![]()
![]()
uni
pressedienst Pressestelle der Universität Zürich
Felix Mäder (fmaeder@zuv.unizh.ch)
http://www.unizh.ch/upd/magazin/1-96/impfstoffe.html
Last update: 1-APR-96